Much of our work is concerned with LIN-12/Notch, the receptor component of one of the major signaling systems for specifying cell fate during animal development. Mutations in core components and modulators of the LIN-12/Notch pathway have been implicated in cancer, Alzheimer's disease, and other diseases and syndromes. Using C. elegans , we study how LIN-12/Notch signaling is modulated during normal development and identify potential ways it may be modulated to combat disease.
In our developmental studies, we aim to understand the fundamental logic and molecular events that govern cell fate decision-making. We mainly study two simple cell fate decision paradigms in reproductive system development: the "AC/VU decision" and "VPC fate specification" during vulval induction. These simple paradigms offer the opportunity to apply powerful methods of genetic analysis to fundamental mechanisms of cell fate specification that operate in all animals. During these events, different modulatory mechanisms regulate LIN-12/Notch activity and different signaling inputs are integrated so that correct cell fate decisions are made. We are continuing to make advances in elucidating these mechanisms.
More recently, we have been studying "global mechanisms" for coordinating gonadogenesis and vulval induction with developmental progression of other body parts. We have found that PTEN and TOR, both conserved regulators that mediate global responses in mammals, act nonautonomously to regulate reproductive system development. We aim to identify the modes and mechanisms of communication that mediate this interorgan communication.
All of our work exploits the "awesome power" of C. elegans genetics. In particular, we use sensitive and specific suppressor and enhancer screens, genetic mosaic analysis, and genome engineering methods to manipulate and visualize endogenous genes and their products. These tools, combined with genetically-encoded signaling biosensors we have developed, long-term imaging using microfluidic devices, and cell-specific transcriptome profiling will allow us to gain unprecedented insight and resolution into signaling regulation and integration.
LIN-12/Notch signaling
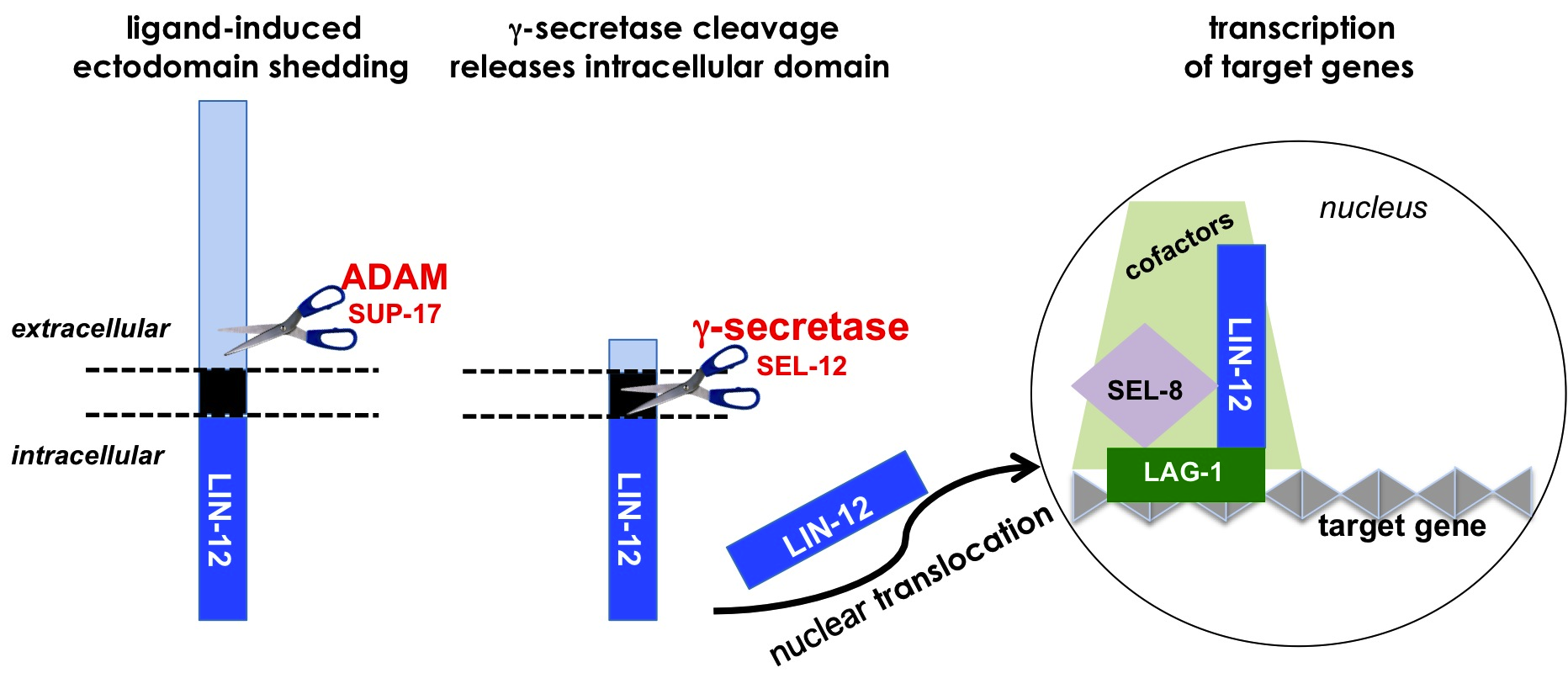
The main events in signal transduction have been elucidated:
A ligand presented by a neighboring cell binds to the ectodomain of LIN-12, activating signal transduction by exposing a cleavage site in the ectodomain.
An ADAM protease such as SUP-17/ADAM10 mediates cleavage at this site, resulting in shedding of the ectodomain.
The resulting transmembrane stub is cleaved within the transmembrane domain by a multiprotein enzyme called "γ-secretase," which contains the catalytic subunit SEL-12/Presenilin.
The intracellular domain, thereby released from its tether, translocates to the nucleus.
The intracellular domain, in a complex with a sequence-specific DNA binding protein, promotes target gene expression.
Genetic analysis was crucial for elucidating the mechanism of signal tranduction and is now leading to deeper understanding of how signaling is modulated. In addition, this basic signaling mechanism enables strategies for identifying target genes and for identifying and analyzing new factors that influence signal transduction, and recently, we have developed tools that facilitate manipulation of LIN-12 and quantitation of signal transduction in vivo.
We recently discovered a surprising evolutionary plasticity in the mechanism by which ligand activates Notch in C. elegans . In Drosophila and mammals, Epsin/Clathrin-mediated endocytosis of ligand in the signaling cell generates a "pulling force" on Notch to expose the ADAM cleavage site. However, we found that neither Epsin nor the intracellular domain of canonical transmembrane ligands are required for ligand function in C. elegans , and that "less force" is needed to expose the ADAM cleavage site of C. elegans Notch proteins because they lack a specific structural element found in other Notch proteins.
Greenwald, I. (2012) Notch and the awesome power of genetics. GeneticsPMID: 22785620 Greenwald, I. and Kovall, R. (2013) Notch signaling: genetics and structure. Wormbook. Langridge, P.D., Garcia-Diaz, A., Chan, J.Y. Greenwald, I. and Struhl, G. (2022). Evolutionary plasticity in the requirement for force exerted by ligand endocytosis to activate C. elegans Notch proteins. Curr. BiolPMID: 35349791 Shaffer, J.M. and Greenwald, I. (2022a) Floxed exon (Flexon): A flexibly positioned stop cassette for recombinase-mediated conditional gene expression. PNASPMID: 35027456 Shaffer, J.M. and Greenwald, I. (2022b) SALSA, a genetically encoded biosensor for spatiotemporal quantification of Notch signal transduction in vivo. Dev. CellPMID: 35413239
AC/VU decision
Stochastic mechanisms and feedback mechanisms during lateral specification
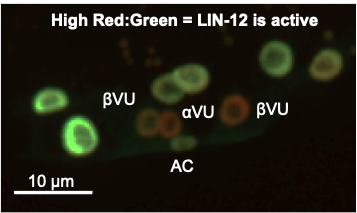
α cells express lin-12 and lag-2. Feedback causes positive autoregulation of lin-12 transcription and degradation of HLH-2 to downregulate lag-2 transcription in the presumptive VU.A Notch biosensor demonstrates high LIN-12 activity in VUs.
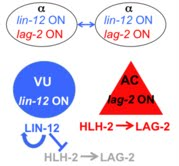
The ventral uterus of hermaphrodites is derived from four cells of the somatic gonad primordium, two "α" cells and their sisters, the "β" cells. The two α cells undergo the "AC/VU decision", a simple paradigm for lateral specification. The β cells are the sisters of the a cells; they lose the potential to be the AC early and generally adopt a VU fate even in a lin-12 null mutant.
During the AC/VU decision, the two α cells communicate so that only one AC is made. Both cells initially express lin-12 and lag-2, the gene encoding its ligand for this decision. As the decision progresses, a stochastic difference in lin-12 acticity is amplifies by feedback mechanisms such that the transcription of both genes becomes mutually exclusive in each cell. The α cell that has the "edge" in LIN-12 activation continues to express lin-12 and becomes the VU; the other α cell becomes the AC and continues to express lag-2. HLH-2, the sole E transcription factor ortholog is initially expressed in the two α and two β cells, endowing them with the potential to become the AC, and then becomes degraded in the presumptive VU in response to LIN-12 activation by a novel dimerization-dependent mechanism.
Using high-throughput lineage analysis in a microfluidic device, CRISPR/Cas9-generated, fluorescently-tagged endogenous genes, and a novel Notch activity sensor, "SALSA," we have gained new insight into this edge develops. We discovered that the main stochastic events that lead to a difference in lin-12 activity occur prior to the birth of the α cells. A highly predictive event is that the first parent of an α cell (Z1.pp or Z4.aa) to express HLH-2, which biases its α daughter to become the αVU. Because the onset of HLH-2 expression in the parents is linked to their birth, and HLH-2 is initially required for transcription of lin-12, earlier expression of HLH-2 in one parent gives its daughter α cell a head start toward the onset of LIN-12 activation by ligand from neighboring cells.
Current projects in the lab aim to achieve a deeper mechanistic understanding of this paradigmatic cell fate decision. We are also interested in certain evo-devo aspects of gonadogenesis that we hope to pursue in the future.
Sallee, M.D. and Greenwald, I. (2015) Dimerization-driven degradation of C. elegans and human E proteins. Genes Dev.PMID: 26159995 Sallee, M.D.*, Littleford, H.E.* and Greenwald, I. (2017) A bhlh code for sexually dimorphic form and function of the C. elegans somatic gonad. Curr. Biol.PMID: 28602651 Attner, M.A., Keil, W., Benavidez, J.M. and Greenwald, I. (2019) HLH-2/E2A expression links stochastic and deterministic elements of a cell fate decision during C. elegans gonadogenesis. Curr. Biol.PMID: 31402303 Littleford, H.E.*, Kiontke, K.*, Fitch, D.H.A.+, and Greenwald I+. (2021) hlh-12, a gene that is necessary and sufficient to promote migration of gonadal regulatory cells in Caenorhabditis elegans, evolved within the Caenorhabditis clade. GeneticsPMID: 34740245 Benavidez, J.M., Kim, J.H. and Greenwald, I. (2022) Influences of HLH-2 stability on anchor cell fate specification during Caenorhabditis elegans gonadogenesis. G3PMID: 35134193 Shaffer, J.M. and Greenwald, I. (2022b) SALSA, a genetically encoded biosensor for spatiotemporal quantification of Notch signal transduction in vivo. Dev. Cell.PMID: 35413239
Regulating and integrating LIN-12/Notch and EGFR-Ras-ERK signaling
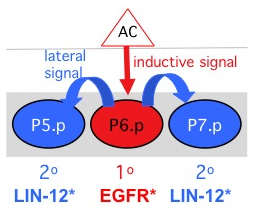
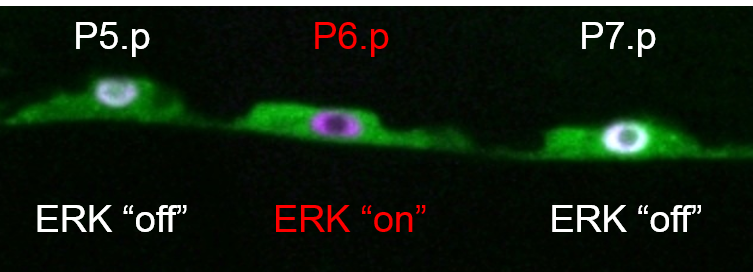
Spatial Patterning of vulvas fates.ERK activity in P6.p revealed using the biosensor ERK-nKTR.LIN-12/Notch activity in P5.p and P7.p descendants revealed using a Notch biosensor.
Six VPCs, named P3.p-P8.p, have the potential to generate vulval cells. In the L3 stage, the EGF-like inductive signal produced by the anchor cell of the gonad activates EGFR-Ras-ERK signaling in P6.p, causing it to adopt the "1o fate" and to produce a "lateral signal". The lateral signal consists of ligands for LIN-12/Notch, which activate LIN-12 in the neighboring cells, P5.p and P7.p, specifying them to adopt the "2o fate". The descendants of the 1o and 2o VPCs undergo morphogenesis to become the vulva.
This invariant and correct pattern of vulval fates depends on proper spatial and temporal control of these signaling pathways. We have identified multiple mechanisms that regulate EGFR and LIN-12/Notch signaling activity and crosstalk between them to achieve the correct pattern of VPC fates. For these studies we have developed tools for quantitative and dynamic studies of signaling in live worms, including genetically-encoded biosensors for ERK and LIN-12/Notch activity.
de la Cova, C., Townley, R., Regot, S.* and Greenwald, I.* (2017) A real-time biosensor for ERK activity reveals signaling dynamics during C. elegans cell fate specification. Dev CellPMID: 28826819Underwood, R.S., Deng, Y. and Greenwald, I. (2017) Integration of EGFR and LIN-12/Notch signaling by LIN-1/Elk1, the Cdk8 kinase module, and SUR-2/Med23 in Vulval Precursor Cell fate patterning in Caenorhabditis elegans.GeneticsPMID: 28954762Deng Y., Luo K.L., Shaye D.D., Greenwald I. (2019) A Screen of the Conserved Kinome for Negative Regulators of LIN-12 Negative Regulatory Region ("NRR")-Missense Activity in Caenorhabditis elegans.G3 PMID: 31519743 de la Cova C., Townley R., Greenwald I. (2020) Negative feedback by conserved kinases patterns the degradation of Caenorhabditis elegans Raf in vulval fate patterning. DevelopmentPMID: 33144396 Luo K.L., Underwood R.S., Greenwald I. (2020) Positive autoregulation of lag-1 in response to LIN-12 activation in cell fate decisions during C. elegans reproductive system development. DevelopmentPMID: 32839181
Global regulation of gonad and vulva developmental progression
Dauer larvae and other arrest paradigms
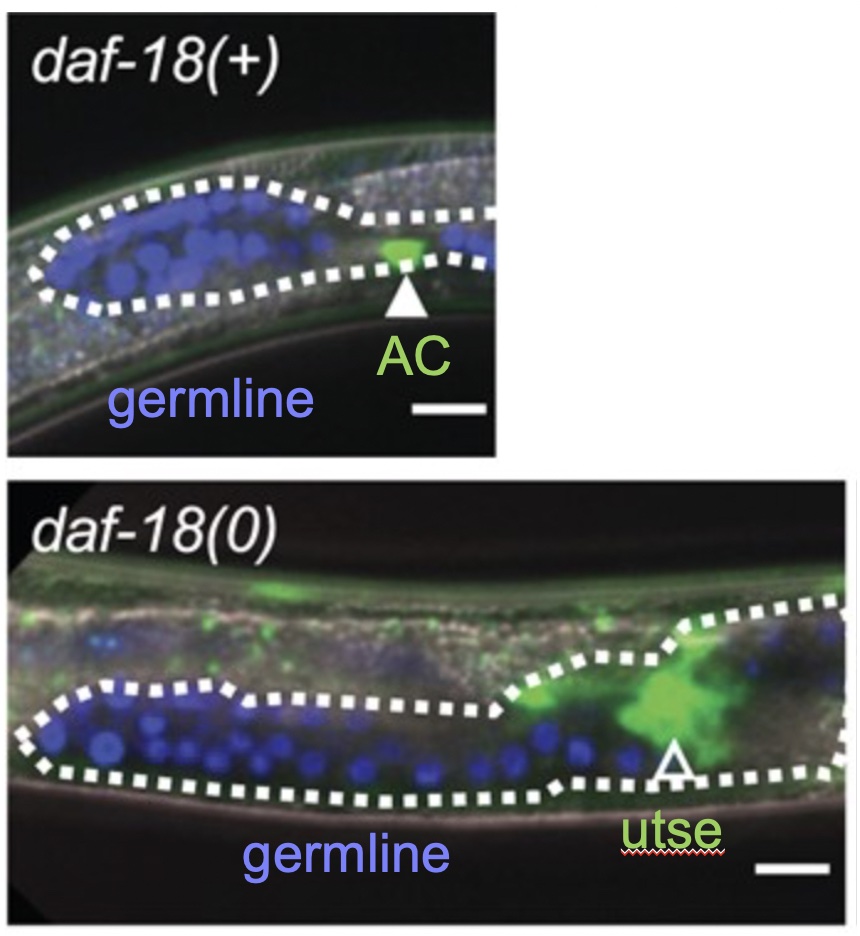
Top, dauer larva showing suspended development of the somatic gonad and germline. Bottom, loss of daf-18/PTEN leads to developmental progression of both the somatic gonad and germline.
During animal development, development requires coordinated developmental progression of all body parts. In C. elegans, such global mechanisms regulate somatic gonad, germline, and vulval progenitors, and utilize conserved signaling pathways that also mediate global responses in mammals.
C. elegans larvae display remarkable developmental plasticity in response to environmental conditions. In favorable conditions, C. elegans progresses rapidly and continuously through four larval stages (L1-L4) to reproductive adulthood. However, starvation or other adverse conditions causes global suspension of development at different points. The most dramatic example is the dauer larva, a state of diapause in which robust mechanisms to maintain prolonged cellular quiescence and developmental potential of progenitor cells are engaged. Dauer entry suspends gonadogenesis and vulval induction, and we have found that DAF-18/PTEN acts nonautonomously to maintain reproductive system precursor cell quiescence in dauer larvae.
We aim to identify the modes and mechanisms of communication that mediate this interorgan communication.
Karp, X. and Greenwald, I. (2013) Control of cell fate plasticity and maintenance of multipotency by DAF-16/FoxO in quiescent C. elegans. Proc. Natl. Acad. Sci.PMID: 233416331 Tenen, C.C. and Greenwald, I. (2019) Cell non-autonomous function of daf-18/PTEN in the somatic gonad coordinates somatic gonad and germline development in C. elegans dauer larvae. Curr. Biol.PMID: 30827916 Wittes, J. and Greenwald, I. (2022) Genetic analysis of DAF-18/PTEN missense mutants for the ability to maintain quiescence of the somatic gonad and germ line in Caenorhabditis elegans dauer larvae. G3(Bethesda)PMID: 35451467 O'Keeffe, C. and Greenwald, I. (2022) EGFR signal transduction is downregulated in C. elegans vulval precursor cells during dauer diapause. DevelopmentPMID: 36227589